8
Kolme keskeistä aivojen muovautuvuuteen liittyvää ja oppimista tukevaa mekanismia ovat synaptogeneesi (engl. synaptogenesis), pitkäkestoinen potentiaatio (engl. long-term potentiation, LTP) ja myelinisaatio (engl. myelination).
Synaptogeneesillä tarkoitetaan hermosolujen välisten yhteyksien, eli synapsien, syntymistä. Yhtäaikaisesti aktivoituvat hermosolut muodostavat ja vahvistavat välillään olevia synapseja. “Muistijälkien muodostuessa niitä säilyttävien solujen välisissä synapseissa ilmenee hebbiläistä muovautuvuutta” (Kim, Choi & Kaang, 2018). On esitetty, että etenkin synapsien tasolla tapahtuva yhteyksien vahvistuminen ja toisaalta taas toiminnan estyminen ovat olennaisia tekijöitä muistijälkien muodostumisessa (Takeuchi, Duszkiewicz & Morris, 2014).
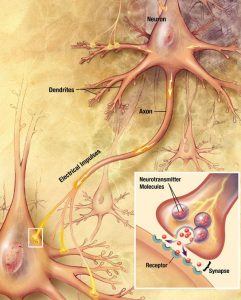
Pitkäkestoisella potentiaatio (LTP) tarkoitetaan yhteyksien muuttumista pysyvämmiksi. Oppiminen näkyy muutoksissa hermosolujen välisissä yhteyksissä, mutta kaikki yhteydet eivät ole samanlaisia. Osa yhteyksistä on nopeita ja väliaikaisia, ja ne tukevat lyhytkestoista musitia. Toiset ovat kestävämpiä ja voivat säilyä tuntien, päivien, tai jopa elämän ajan (ts. säilömuisti). Kestävän oppimisen kannalta on hyödyllisintä keskittyä siihen, miten pitkäkestoiset yhteydet muodostuvat. Pitkäkestoisella potentiaatiolla uskotaan olevan tässä avainasema. Potentiaation aikana luodaan uusia proteiineja, minkä on havaittu käynnistyvän tietynlaisen aktivaation seurauksena, kuten reseptissä (Fields, 2005). Niin eläintutkimuksissa kuin hermosoluja petrimaljoissa tarkastelevissa tutkimuksissa havaittiin potentiaation käynnistävä 3 x 2 -kaava: kolme “aktivaatiota”, joiden välissä oli kaksi 10 minuutin taukoa. Erityisen tärkeää tutkimuksessa oli se, että tauot olivat ainakin 10 minuutin pituisia, sillä tällöin aivojen ajatellaan käsittelevän aktivaatioita erillisinä kokemuksina. Lisäksi useammin toistuvien kokemusten muistijälki lujittuu todennäköisemmin. Näin ollen 3 x 2 eli kolme saman verkoston (muistijäljen) aktivaatiota aivoissa 10 minuutin välein oli vähimmäisvaatimus pitkäkestoiselle potentiaatiolle ja siten muistijäljen lujittumiselle säilömuistiin. Tähän tutkimustietoon perustuu myös ’Jaksotettu oppiminen’-opetusstrategia (Kelley & Whatson, 2013).
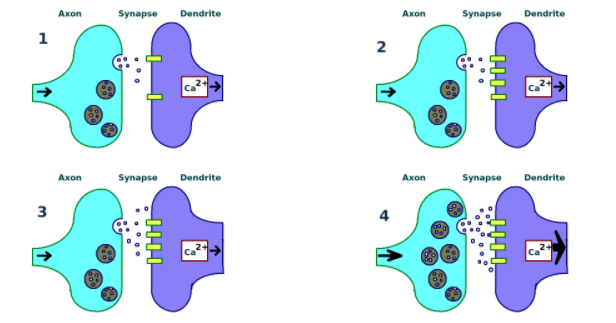
(1) A synapse is repeatedly stimulated. “A synapse is repeatedly stimulated, sending neurotransmitters from the axon terminal (left) across the synapse to the dendrites of a second neuron (right).” (2) More receptors are found on the dendrite. (3) More neurotransmitters are produced. (4) A stronger link between neurons is established. “Final stage of long term potentiation. Two nerves (left and right) are linked with a strong signal which works quickly. Scientists believe long term potentiation is the primary way that the brain learns.” (Source: Wikipedia, Creator: Tomwsulcer)
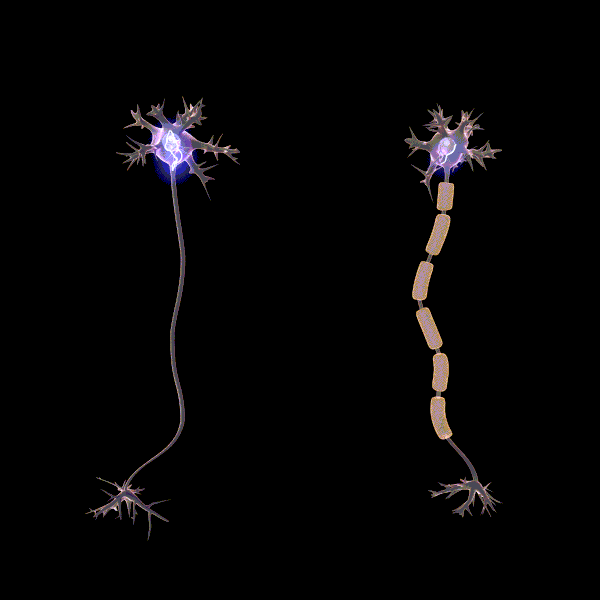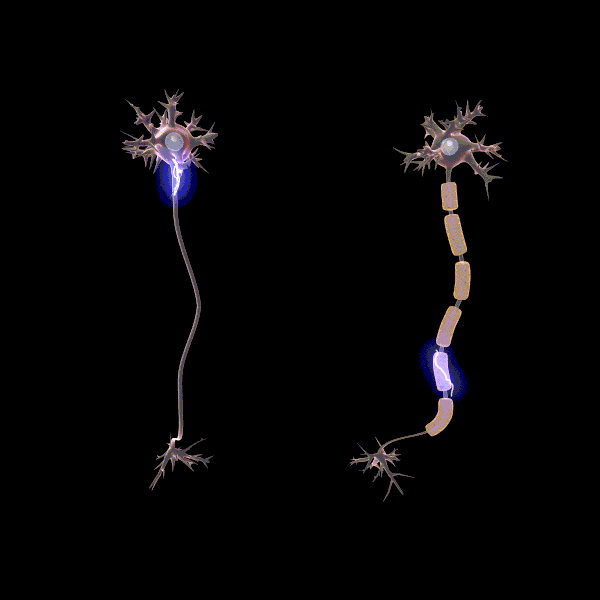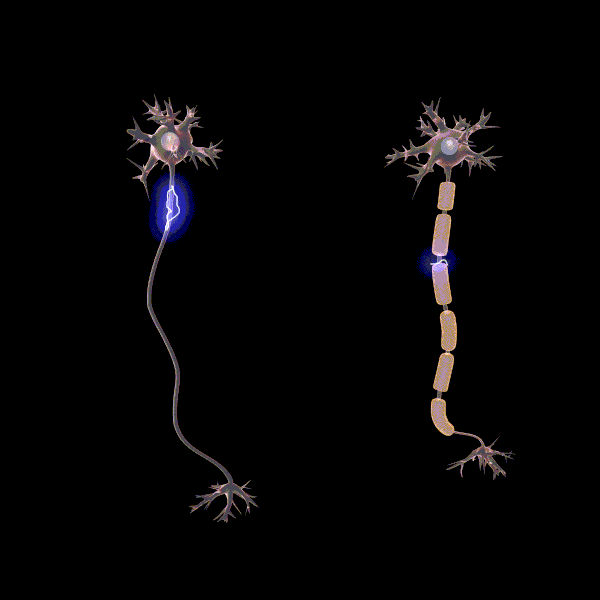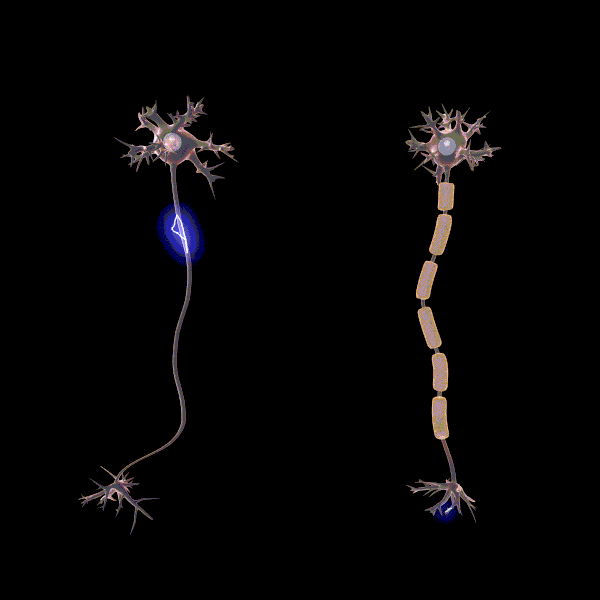
Myelinisaatiolla viitataan yhteyksien tehostamiseen ja myelinisaation tiedetäänkin lisäävän sähkön johtumisen nopeutta yksittäisissä aksoneissa, eli hermosolun viejähaarakkeissa (Pan ym., 2020). Myeliinin rooli oppimisessa ei ole täysin selvä, mutta erään teorian mukaan myelinisaatio mahdollistaa usein käytettyjen verkostojen tehostamisen siten, että ne käyttävät vähemmän resursseja. Esimerkiksi myeliinituppi muodostuu aksonin ympärille eristäen sen, jolloin energiaa menee vähemmän hukkaan hermosolun aktivoituessa. “Myelinisaatio on mahdollisesti hebbiläisen muovautuvuuden tapaan tiettyjen yhteyksien vahvistamista ja edistää viestintää hajautettujen hermoverkkokokonaisuuksien välillä” (Pan ym., 2020).
Lähteet: Osa 2
- Goelet, P., Castellucci, V. F., Schacher, S., & Kandel, E. R. (1986). The long and the short of long–term memory—a molecular framework. Nature, 322(6078), 419.
- Sutton, M. A., & Schuman, E. M. (2006). Dendritic protein synthesis, synaptic plasticity, and memory. Cell, 127(1), 49-58.
- *Farely, P. (2020 February 10). Long-Term Learning Requires New Nerve Insulation. Retrieved from https://www.ucsf.edu/news/2020/02/416621/long-term-learning-requires-new-nerve-insulation
- Kim, J. I., Choi, D. I., & Kaang, B. K. (2018). Strengthened connections between engrams encode specific memories. BMB reports, 51(8), 369.
- Takeuchi, T., Duszkiewicz, A. J., & Morris, R. G. (2014). The synaptic plasticity and memory hypothesis: encoding, storage and persistence. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1633), 20130288.
- Fields, R. D. (2005). Making memories stick. Scientific American, 292(2), 74-81.
- Kelley, P., & Whatson, T. (2013). Making long-term memories in minutes: a spaced learning pattern from memory research in education. Frontiers in human neuroscience, 7, 589.
- Davis, R. L., & Zhong, Y. (2017). The biology of forgetting—a perspective. Neuron, 95(3), 490-503.
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., & Kheirbek, M. A. (2020). Preservation of a remote fear memory requires new myelin formation. Nature neuroscience, 23(4), 487-499.
- Farely, P. (2020 February 10). Long-Term Learning Requires New Nerve Insulation. Retrieved from https://www.ucsf.edu/news/2020/02/416621/long-term-learning-requires-new-nerve-insulation

{kind=link}
{kind=link}